Børsteorme (Polychaeta) er ledorme og findes i de interstitielle miljøer nede i havbundens sedimenter. De udgør en blandet gruppe, som tidligere blev regnet for meget primitiv og anbragt i en særlig gruppe, „Archiannelida“. Men de er ikke primitive, tværtimod er der tale om stærkt specialicerede børsteorme. De er sekundært reducerede og er – uafhængigt af hinanden – opstået fra forskellige grupper af børsteorme.
Som eksempel kan nævnes, at arter i den „primitive“ familie Dinophilidae, som omfatter de allermindste arter i meiofaunaen, i virkeligheden nedstammer fra den mere avancerede familie Dorvilleidae. Man har nemlig inden for de sidste få år fundet overgangsformer fra store dorvilleider med kæber og komplicerede børstevedhæng til små former uden kæber, men med ciliebånd, der ellers kun findes hos larverne. Slægten Dinophilus er en sådan reduceret form uden kæber; den er meget almindelig i danske farvande. Hanner af dinophilider bliver ikke meget større end 0,1 mm. Da dyrene er helt beklædt med cilier, kan de let forveksles med fladorme.
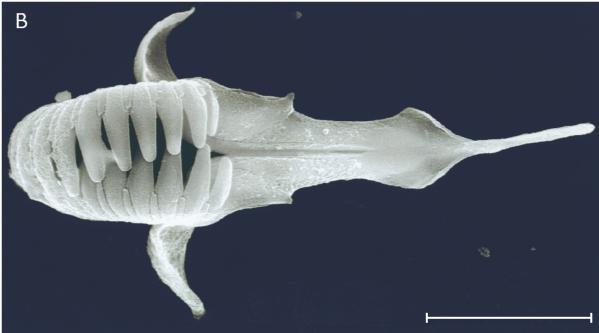
Senest har danske forskere fundet den mærkelige slægt Neotenotrocha, som måske er de mindste børsteorme i verden. Dyrene har kæber, men ellers ligner de fuldstændig en larve af en dorvilleid med de karakteristiske ciliebånd. Alligevel har dyrene kønsorganer (figur 6-16).
Det interstitielle miljø har ligefrem været et „rugekammer“ for en evolutionær eksplosion af nye former af små børsteorme, men de er ikke alle så reducerede som Neotenotrocha. Repræsentanter for familien Nerillidae har bevaret deres palper, børster og parapodier og har dertil specielle tilpasninger til livet mellem sandskornene. Arten Nerillidium troglochaetoides har yngelpleje (figur 6-14 og 27). Ægget er fastklæbet til bagenden, og afkommet udvikler sig her. Larvestadiet mangler som hos mange andre interstitielle dyr.
De interstitielle børsteorme er normalt sjældne, dvs. at man kun finder nogle få eksemplarer i hver prøve. En undtagelse er dog arter af slægten Polygordius. Den kan i grove sedimenter som skalgrus eller sand med skalfragmenter forekomme i massevis. Man har ligefrem opkaldt denne sedimenttype, Polygordius--sand, efter denne slægt.
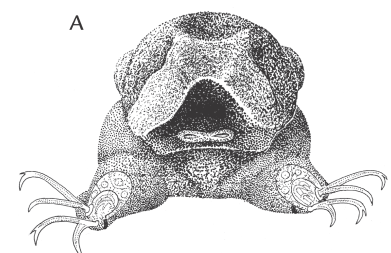
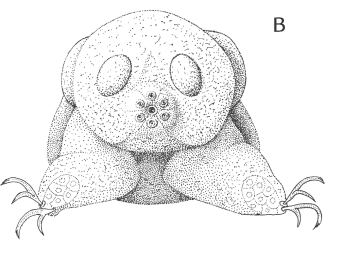
De interstitielle børsteorme kan være så specialiserede, at de kan være svære at erkende som ledorme. Det gælder for familien Psammodrilidae, som har to arter i Danmark. Den store art, Psammodrilus balanoglossoides, er ret almindelig ved Ellekilde Hage i Øresund, hvor den lever i tynde rør af sammenkittet sand. Man kan ikke rigtigt kalde den for en interstitiel børsteorm, da den er for stor til at bevæge sig mellem sandskornene. Alligevel er den helt beklædt med cilier ligesom mange små interstitielle dyr. Den lille art, Psammodriloides fauveli, lever derimod i skalgrus og er altså fuldgyldigt interstitiel. Den er til gengæld stærkt reduceret med korte børstevedhæng og mangler krave (figur 6-17).





Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.