

Fotosyntese er en af de vigtigste biologiske processer på Jorden, da den producerer det meste af atmosfærens oxygen (ilt), som næsten alt liv afhænger af. Fotosyntesen foregår især i grønne planter, alger og visse bakterier.

Fotosyntese er af altafgørende betydning for opretholdelsen af Jordens stofkredsløb.
Formlen for fotosyntese
I fotosyntesen omdannes kuldioxid og vand ved hjælp af sollysets energi til oxygen og organisk stof i form af glukose (druesukker). Glukose omdannes siden til andre næringsstoffer, der bruges af planterne selv og af dyr.
Formlen for fotosyntesen skrives ofte sådan:
kuldioxid + vand + sollys → glukose + oxygen
eller mere præcist:
6 CO2 + 6 H2O + sollys → C6H12O6 + 6 O2
Stofkredsløb og energi

Kulstofkredsløbet drives af fotosyntesen ved dens opbygning af kulhydrater samt af forbrænding og afgivelse af kuldioxid, som er tilgængelig for planterne. En stor mængde plantemateriale er tidligere i Jordens historie blevet begravet og senere omdannet til de olie- og gasforekomster, hvis kemiske energi i dag udnyttes som fossilt brændsel; fra disse afgives CO2til atmosfæren, hvor CO2-andelen er steget fra 310 ppm i 1960 til 420 ppm i 2020.
Fotosyntese er af altafgørende betydning for opretholdelsen af Jordens stofkredsløb. Ved fotosyntese dannes årligt ca. 200 milliarder tons organiske forbindelser. Det svarer til, at al atmosfærens kuldioxid i løbet af en periode på 300 år omdannes til organisk stof, og at al atmosfærens oxygen fornyes i løbet af 2000 år. Når atmosfærens indhold af kuldioxid og oxygen (hhv. 0,042 % og 21 %) alligevel holder sig næsten konstant, skyldes det, at alle levende organismer, der ikke kan udføre fotosyntese, kræver energi tilført i form af næringsstoffer for at kunne opretholde livsfunktionerne. Næringsstofferne er typisk de organiske forbindelser, der dannes ved fotosyntese, og energien frigives ved nedbrydning til kuldioxid og vand under forbrug af oxygen ved ånding, respiration (se respiration hos planter). Også planter og alger nedbryder konstant organiske forbindelser ved respiration, men i lys overstiger fotosyntese respirationen, således at der finder en nettostofproduktion sted.
Dyr kan ikke bruge sollys som en direkte energikilde og skal i stedet altid have de nødvendige organiske forbindelser tilført ved at æde planter eller andre dyr, som har fået deres energibehov dækket ved at æde planter. Visse dyr og svampe (laver) får dog tilført næring ved at leve i symbiose med alger, som udnytter lysenergien.
De fossile brændstoffer, kul, olie og naturgas, som vi forbruger i vores hverdag, er dannet ved nedbrydning af dyr og planter, som for millioner af år siden direkte eller indirekte udnyttede Solen som energikilde. Næsten al energi til organisk stofproduktion kommer således i sidste ende fra sollyset, og de produkter, der dannes ved fotosyntesen, forbruges senere hen ved respirationen. På denne måde kommer Jordens kulstofkredsløb i balance. En af de få og kvantitativt ubetydelige undtagelser er den bakterielle stofproduktion, der foregår ved udnyttelse af den kemisk bundne energi i stoffer, som bliver frigivet ved varme, undersøiske kilder, der ligger i absolut mørke
Fotosyntese hos planter

Fotosyntese hos planter ved lys af forskellig bølgelængde. Fotosyntesen er lavest ved ultraviolet, grønt og infrarødt lys og højest ved blåt eller rødt lys.
Hovedparten af den fotosyntetiske produktion finder sted i højere planter. Planter binder normalt 2-5 % af den solenergi, der rammer dem, i organisk stof. Resten går tabt. Hos planter finder fotosyntesen helt overvejende sted i bladene.
Bladet
Et typisk blad består af bladkød (mesofylceller), karstrenge og epidermis. I bladkødet findes størsteparten af bladets kloroplaster, grønkorn; det er i disse, at fotosyntesen finder sted. Karstrengene er plantens transportsystem for vand, salte og organiske forbindelser, herunder kulhydrater dannet ved fotosyntesen.
Epidermis fungerer som bladets beskyttende overflade og indeholder kun få kloroplaster, men er alligevel af stor betydning for fotosyntesen, idet det er her, bladets spalteåbninger (stomata) findes. Ved åbning og lukning af spalteåbningerne styrer planten dels sin vandhusholdning, dels det for de fotosyntetiske processer betydningsfulde optag af kuldioxid fra den omgivende luft.
Kloroplaster

Principtegning af en kloroplast fra en frøplante. Yderst ses kloroplastens dobbeltmembran (ydre og indre membran), der omgiver stroma med grana- og stromathylakoider, hvis membraner afgrænser hulrum, der er i indbyrdes kontakt. Hos planter, der er tilpasset skyggefulde områder, er thylakoidstablen i grana højere end hos planter tilpasset stærkt sollys.
Kloroplaster er typisk ellipseformede legemer med en længde på ca. 5-10 micrometer (0,005- 0,01 mm). I deres indre findes et membransystem, thylakoidmembranerne, opbygget af delvis sammentrykkede membransække, der er ordnet i stabler, grana, som er indbyrdes forbundne ved hjælp af enkelte membransække, stromalameller.
Alle plantens fotosyntetiske pigmenter (klorofyl a og b samt carotenoider såsom β-caroten og xanthofyller) er bundet til proteiner placeret i kloroplasternes indre membraner. Det er pigmenterne i disse membraner, der sætter planter i stand til at indfange (absorbere) lys. De fotosyntetiske pigmenter absorberer først og fremmest lys med bølgelængder svarende til blåt og rødt lys. Lys i det grønne bølgelængdeområde kastes tilbage eller passerer uhindret igennem bladet, og det er derfor, at planter ser grønne ud.
Lysreaktionerne
De fotosyntetiske pigmenter er bundet til forskellige proteiner i membranen, som er organiseret i to forskellige funktionelle enheder, kaldet fotosystem I og II. Hvert fotosystem indeholder 250-400 pigmentmolekyler. Hvert enkelt pigmentmolekyle i en sådan enhed kan absorbere lys, hvorved det bliver anslået, dvs. at en af dets elektroner bliver bragt op på et højere energiniveau. Pigmentmolekylerne er pga. de proteiner, de er bundet til, placeret på en sådan måde i forhold til hinanden, at den lysenergi, der er absorberet af et pigmentmolekyle, effektivt kan overføres til et andet. Derved kommer de til at fungere som et samlet antennesystem, der kan indfange lysenergi.
I hvert fotosystem findes et specielt par klorofyl a-molekyler, reaktionscentret. Når lysenergien overføres hertil, og reaktionscentret således bliver anslået, overføres en elektron fra et af reaktionscentrets klorofylmolekyler til et elektronacceptormolekyle. Herved bliver elektronacceptoren reduceret (negativt ladet), og reaktionscentret oxideret (positivt ladet).
De proteiner, som de to fotosystemer er opbygget af, har yderligere en række elektronacceptorer og -donorer bundet til sig. Disse muliggør dels hurtig videretransport af den elektron, der frigives fra reaktionscentret, dels transport af elektroner til det oxiderede reaktionscenter, der derved bringes tilbage til udgangstilstanden og således kan indgå i en ny reaktion. De to fotosystemer katalyserer således hver især transport af elektroner.
De to fotosystemer er funktionsmæssigt koblede, således at fotosystem II leverer elektroner til fotosystem I. Fotosystem II er overvejende placeret i grana, mens fotosystem I er placeret i stromalamellerne. Samlet fører elektrontransporten i fotosystem II til oxidation af vand med frigivelse af ilt, mens elektrontransporten i fotosystem I fører til reduktion af stoffet NADP+ til det energirige NADPH.
De elektronacceptorer og -donorer, som medvirker i elektrontransporten, er alle bundet til proteiner, der sørger for, at acceptorerne er placeret optimalt i forhold til hinanden. De hurtigste elektronoverførsler foregår således i løbet af 3 picosekunder (3 billiontedele sekund), de langsomste i løbet af ca. 0,02 sekund. Blandt andet kinoner, jern-svovl-centre og cytochromer indgår som transportører af elektroner.
ATP-dannelse
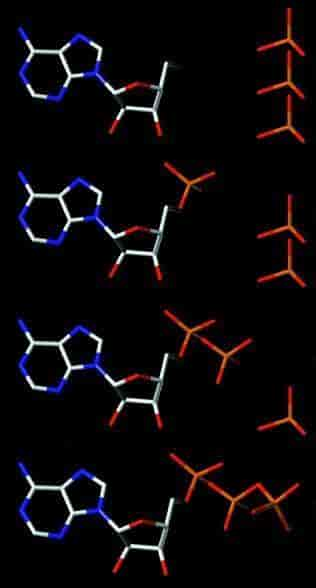
ATP (nederst) er sammensat af adenosin og tre fosfatgrupper (de røde og orange strukturer til højre). Fosfat kan spaltes fra i tre trin (nederst til øverst), men det er kun ved de to første, der frigives energi. I kroppen er spaltningen af ATP til ADP (adenosindifosfat) og fosfat langt den almindeligste.
I både fotosystem I og II transporteres elektroner fra thylakoid-membransækkens indre til dens yderside. Denne elektrontransport fører samtidig til en ophobning af hydrogenioner (brintioner, H+) i membransækkens indre. Den koncentrationsforskel, der herved opbygges, udnyttes af et enzym, en ATP-syntase, der ligeledes er placeret i membranen, til dannelse af ATP. Denne proces kaldes fotofosforylering (se også oxidativ fosforylering, som er en lignende delprocess i respirationen).
ATP er en særdeles energirig forbindelse, og vha. den fotosyntetiske elektrontransport har planten således fået omdannet lysenergi til energirige kemiske forbindelser, der kan bruges, når planten har behov for dem. De processer, der fører til dannelse af NADPH og ATP, kaldes samlet for fotosyntesens lysreaktioner, idet lys er en forudsætning for, at de kan forløbe.
Kulstofreaktionerne
Fotosyntesens lysreaktioner foregår i kloroplasternes indre. De efterfølgende reaktioner, hvorved kuldioxid ved forbrug af ATP og NADPH indbygges i organiske forbindelser, katalyseres af enzymer i kloroplastens stroma, dvs. den opløsning, der afgrænses af kloroplasternes to ydre membraner og det indre membransystem. Disse reaktioner kaldes fotosyntesens kulstofreaktioner eller Calvin-cyklus efter den amerikanske biokemiker Melvin Calvin, der klarlagde dem. Da Calvin-cyklus kun kan foregå, når lysreaktionerne leverer NADPH og ATP, og da flere af enzymerne i Calvin-cyklus aktiveres af lys, er den gamle betegnelse ’mørkereaktioner’ for kulstofreaktionerne misvisende.
Biosyntese og eksport
Luftens kuldioxid kommer til kloroplasternes stroma ved diffusion og her fikseres den af enzymet Rubisco, ribulose-bisfosfat-carboxylase, i en organisk forbindelse med tre kulstofatomer (C3-forbindelser). Denne C3-forbindelse og de forbindelser, den omdannes til i Calvin cyklus, bliver brugt som udgangsmateriale til syntese af mange af de forbindelser som kloroplasten og cellen har brug for, f.eks. sucrose og stivelse. C3-forbindelsen kan også eksporteres til cytosolen, hvor den igen kan indgå i biosyntese, men hvor den også kan bidrage til cellens energiforsyning ved at være substrat for cellens respiration.
Under forhold, hvor Calvin-cyklen forløber så hurtigt, at omdannelsen til sucrose ikke kan følge med, sker der dannelse af stivelse direkte i kloroplastens stroma. Stivelsen nedbrydes så, når der bliver brug for den, fx i mørke.
Via planternes karsystem (floemet) kan overskydende sucrose transporteres fra bladet til andre dele af planten, hvor den enten kan bruges til biosyntese, til energiproduktion i respirationen eller omdannes til oplagsnæring i form af stivelse i fx knolde og frø.
Fotosyntesehastigheden
Den hastighed, hvormed en plante udfører fotosyntese, er afhængig af intensiteten af det lys, der når planten, mængden af kuldioxid, temperaturen, mængden af vand og den omgivende lufts relative fugtighed. Ofte er vandmængden den begrænsende faktor i fotosyntesen. Når planten mangler vand, lukker den sine spalteåbninger (stomata) for at begrænse yderligere tab af vand ved fordampning. Herved begrænses imidlertid også plantens optag af kuldioxid gennem spalteåbningerne og dermed fotosyntesen. Denne begrænsning vil delvis kunne imødegås ved et højere indhold af kuldioxid i atmosfæren. I drivhusgartnerier kan man således opnå en øget produktivitet ved tilførsel af kuldioxid.
Atmosfærens indhold af kuldioxid er lavt. Når et blad modtager fuldt sollys, kan fotosyntesen foregå så hurtigt, at bladet selv med helt åbne spalteåbninger ikke kan optage kuldioxid hurtigt nok fra luften. Ved lavere lysintensiteter bliver respirationen mere og mere betydningsfuld i forhold til fotosyntesen. Den lysintensitet, hvor fotosyntesen og respirationen er lige store, kaldes lyskompensationspunktet og er ca. 2 % af fuldt sollys, bl.a. afhængigt af plantens art.
I en bevoksning vil planternes mange blade delvis skygge for hinanden: De øverste blade vil modtage fuldt sollys, mens de nederste befinder sig i halvskygge. I denne situation kan luftens koncentration af kuldioxid være den begrænsende faktor for fotosyntesen i de øverste bladlag, mens det er mængden af lys, der er begrænsende i de nedre bladlag; samlet for bevoksningen er lyset ofte den begrænsende faktor for den totale fotosyntese.
Adaptationer
Planterne er på mange måder i stand til løbende at tilpasse og optimere deres fotosyntese efter ændringer i omgivelserne. Ændringer i bladtykkelse, bladstilling, kloroplasternes antal, antennesystemernes størrelse samt justering af elektrontransportvejene er nogle eksempler herpå. Nogle planter, fx majs, kan opkoncentrere kuldioxid i nogle af bladkødets celler. Dette kræver energi, men gør planterne i stand til at udnytte luftens kuldioxid bedre. Disse såkaldte C4-planter har en særlig fordel i områder med høj temperatur og begrænsede vandresurser. Mange ørkenplanter, fx kaktus, har kun spalteåbningerne åbne om natten, hvor kuldioxid oplagres i de kødfulde blade og stængler. Om dagen holdes spalteåbningerne lukkede for at hindre for stort vandtab, og fotosyntesen forløber derfor ved brug af den kuldioxid, der blev optaget natten før. Oplagringen af kuldioxid kræver megen energi, og denne form for fotosyntese, CAM-fotosyntese, er kun fordelagtig i meget tørre områder.
Fotosyntese i planteplankton
Umiddelbart forbindes fotosyntese mest med de højere planter, men omkring 40 % af den samlede fotosyntetiske produktion foretages faktisk af planteplankton, fytoplankton, dvs. mikroskopiske alger, der lever i oceanerne. I havet er det normalt mængden af fosfat og nitrat samt andre næringssalte, der er hastighedsbegrænsende faktorer i fotosyntesen. Der kan derfor ved udledning af store mængder næringssalte fra bl.a. landbrug og private husholdninger forekomme kraftig opblomstring af planteplankton. Når planktonet dør, synker det til bunden, hvor det nedbrydes af mikroorganismer under forbrug af oxygen, hvilket kan føre til iltsvind (hypoxia) i de pågældende havområder.
Bakteriel fotosyntese
En række forskellige prokaryote organismer er i stand til at foretage fotosyntese, og det var sandsynligvis hos disse organismer, at mekanismerne bag fotosyntesen oprindelig blev udviklet. Det drejer sig om cyanobakterier og om bakterier i grupperne grønne svovlbakterier (fx Chlorobium), purpursvovlbakterier (fx Thiospirillum), grønne svovlbakterier (fx Chloroflexus) og purpur-ikke-svovlbakterier (fx Rhodospirillum). Hos disse organismer foregår fotosyntesen i pigmentholdige membransække i cellernes cytoplasma. I forbindelse hermed indeholder cyanobakterierne særlige antenne-pigmenter organiseret i såkaldte fykobilisomer.
Hos cyanobakterier er fotosyntesen ligesom hos planter koblet til en spaltning af vand i protoner og oxygen, mens fotosyntesen hos purpur- og grønne svovlbakterier foregår under strengt anaerobe forhold, primært koblet til en spaltning af svovlbrinte (hydrogensulfid, H2S) til protoner og frit svovl. De heterotrofe purpur- og grønne ikke-svovlbakterier kan vokse under tilstedeværelse af ilt, men nogle kan også udføre en anaerob fotosyntese med hydrogen som donor af elektroner.
Hos cyanobakterier og purpurbakterier opbygges det organiske stof gennem Calvin-cyklus efter samme mønster som hos planterne, mens grønne svovlbakterier benytter citronsyrecyklus, der normalt er energiskabende, men her tvinges til at "køre baglæns" og under forbrug af energi opbygge oxaloacetat og citrat. Se også bakterieklorofyl.
Oxygenkatastrofen
Cyanobakterierne opstod i havet for måske mere end 3 mia. år siden. Før den tid havde Jordens atmosfære været oxygenfri (anoxisk), og ophobningen af oxygen (produceret af cyanobakterierne og måske andre fotosyntetiserende organismer) i havene medførte derfor betydelige omvæltninger i økosystemerne. Mange organismer uddøde, da fri oxygen ved sin kemiske reaktivitet under de da herskende forhold var en cellegift, som ødelagde livsvigtige enzymer og nedbrød organiske forbindelser. Samtidig blev det opløste jern i vandmasserne oxideret og udfældedes i båndede jernformationer (se BIF), som i dag brydes flere steder i verden.
Forskning
Fotosyntetiserende bakterier indeholder kun et fotosystem, som er placeret i cellemembranen, samt bakterieklorofyl i stedet for klorofyl. Disse bakterier er på mange måder nemmere at arbejde med end planter, og megen af den viden, vi i dag har om fotosystemernes opbygning, kommer således fra pionerstudier med fotosyntetiserende bakterier. Kendskabet til den nøjagtige placering af elektrontransportørerne i fotosystem II er således baseret på arbejde med purpurbakterier, et arbejde, de tre tyske biokemikere J. Deisenhofer, H. Michel og R. Huber i 1988 tildeltes nobelprisen i kemi for.
Det er håbet, at det udførlige kendskab, vi efterhånden har til fotosystemernes opbygning, vil kunne udnyttes til fremstilling af højeffektive, kunstige solceller, der ligesom planter kan producere kemisk energi ud fra sollys.

Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.