
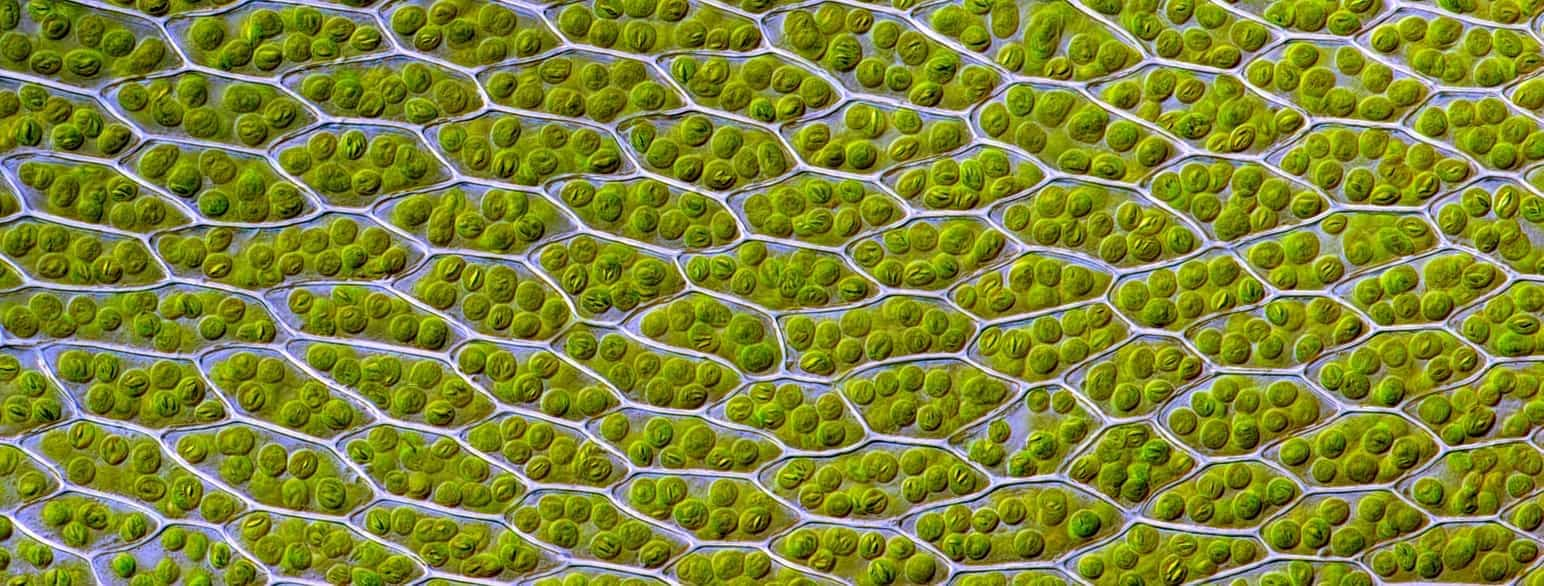
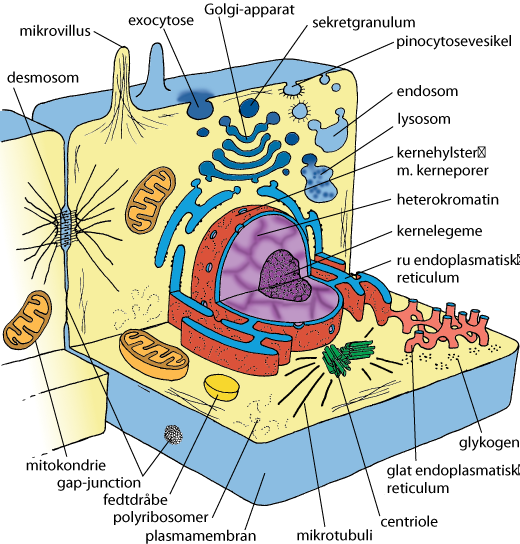


Celle er den mindste enhed for liv. Alle organismer er opbygget af én eller flere celler. Nogle organismer, såsom bakterier, nogle dyr, planter og svampe, består kun af én celle og kaldes encellede organismer. Andre organismer kan bestå af flere identiske celler, der danner kolonier. De fleste organismer består af mange forskellige celler, som samarbejder på forskellige måder. Antallet af celler i disse organismer er normalt enormt, og det afhænger blandt andet af organismens størrelse.
Faktaboks
- Etymologi
-
Ordet celle kommer af latin cella 'kammer', 'aflukke', diminutivformen cellula anvendes ofte i afledninger, fx intracellulær.

{kind=link}
Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.